1.はじめに
南関東では1978-79年、1980-82年、1990-92年、2000-02年、南多摩地域と都心の一部ではさらに2010年にタンポポの在来種・外来種の生育地分布調査が行われました。関西では1973年の兵庫県をを皮切りに、1975年より5年ごとにタンポポの在来種・外来種の分布調査が行われ、やがて西日本の広範な地域で行われ、2020年が5年ごとの調査年になっています。
関東地方平野部に生育する在来種はカントウタンポポ(画像はこちら)とシロバナタンポポ(画像はこちら)です。明治時代はじめに、ヨーロッパから食用にタンポポが輸入されたようです。この子孫が札幌で広がり始めました。当時これを聞いた植物学者の牧野富太郎さんが、「やがて日本中に広がるのではないか」と予言をして、その通りになりました。この外来種を日本ではセイヨウタンポポ(画像はこちら)と呼びました。
2.タンポポの倍数性と生殖方法
日本に持ち込まれたセイヨウタンポポは、染色体(遺伝子の運び屋)を3セット(24本)持っていました(3倍体と呼びます)。日本にもともと生育しているタンポポの中にも、3セットの染色体を持っている種類があります。ちなみに、私たちヒトは一般に両親から1セット(23本)ずつ受け継いで2セット(46本)の染色体を持っています(2倍体と呼びます)。
セイヨウタンポポは、卵細胞や花粉(将来の精核)形成のための細胞分裂(減数分裂)にあたって、染色体を正常に2分できないため、卵細胞形成過程では途中で細胞分裂を止めてしまい、もとの細胞に戻ります。この細胞が受精をせずに細胞分裂を始めて種子(タネ)に育ってしまいます。つまり、親とまったく同じ染色体(遺伝情報)を持った子どもができます。いわゆるクローンです。このように配偶者不要(卵子と精核の融合なし)で子孫ができる生殖方法を、無配(無配偶)生殖、また無融合生殖と呼びます。
一方セイヨウタンポポの花粉形成過程では、二つに分かれる細胞に真半分ずつ染色体を配分することをせずに、いろいろな数の染色体を持った細胞をつくってしまいます。結果的に0本〜24本の染色体を持つ花粉、見た目には大小様々なサイズの花粉がつくられます。これらの花粉の多くは生殖能力を持っていません。ただ、たまたま染色体を2セット(16本)、または3セット(24本)持った花粉がつくられることがあり、この花粉が在来種の2セット(16本)染色体を持った種類のタンポポ(2倍体)に受精すると雑種ができます。この場合、2セットの染色体を持つタンポポの卵細胞は親株の半分に当たる1セット(8本)の染色体を持っているので、受精後は3セット(24本=3倍体)または4セット(32本=4倍体)の染色体を持つ雑種になります。セイヨウタンポポの卵細胞の方は、花が開くころにはすでに種子形成のための細胞分裂が始まっているので、花に正常な花粉が到達しても受精は起こりません。セイヨウタンポポの原産地では、2倍体と3倍体の個体が同じ場所に生えていることがあり、上述したことと同様な子孫形成が行われているとみられますが、染色体は倍体も3倍体も同じ形(遺伝子自体は同じ)なので、雑種の扱いはしていません。
3.受精の有無とメリット、デメリット
受精なしで種子ができるということは、1株あれば無限に子孫をつくれるという計算になり、新天地に少ない、あるいは1個体だけで進出するには都合がよい性質です。しかし、子孫が皆同じ性質を持っているということは、都合の悪い環境変化には抵抗力を持っていないというデメリットもあります。一方、2セットの染色体を持ったタンポポ(2倍体)では、自分の花粉では受精せず、子孫形成には必ず他個体由来の花粉が必要です。これを自家不和合と呼び、近親交配を避ける仕組みになっていて、タンポポで顕著な性質です。こちらは、少数個体では子孫維持がしにくいというデメリットがあります。種子形成に多くの個体が群れて(近くに生育して)いる必要があるのは、デメリットではありますが、一つ一つの個体は性質の異なる遺伝子を保有する結果になり、集団としては環境変化に対応可能というメリットもあります。
4.セイヨウタンポポによる在来種タンポポ駆逐説の風評拡大
ではなぜ、セイヨウタンポポが日本中に広がったのでしょうか?広がったということだけは定性的に(感覚的に)とらえられていましたが、このことを専門的に研究している人がいませんでした。そこで1970年代以降、広がったセイヨウタンポポと目立たなくなった在来種タンポポという二つの現象を結びつけて説明するのに都合の良い見聞が集められ、それらを繋げて、「強いセイヨウタンポポが弱い在来種タンポポを駆逐した」という風説がマスコミを通して広がりました。例えば、セイヨウタンポポは花が多い、だから種子も多い、発芽がよく、早い、成長が早い、体が大きい、タネが軽く、ついている綿毛(冠毛と呼びます)でより遠くまで飛ぶ、セイヨウタンポポはアレロパシーという生育阻害物質を出して周りの草を枯らしているなどを理由としました。それで計算上、数年後にはセイヨウタンポポだらけになり、在来種タンポポより早く生育地を覆ってしまうというのです。この筋書きは日本ではあまり抵抗もなく受け入れられました。その背景には、弱肉強食、優勝劣敗(優者必勝劣者必敗)、生存競争といった、今から振り返ると誤った自然観が教え込まれてきたことがあると思われます。こうした考え方はダーウィンの「種の起源」によって世に出たのですが、ダーウィン自身、数年後に誤りを認めて適者生存という用語に訂正しています。日本で誤ったまま使われ続けたのは、明治時代以降の富国強兵の国造りにとって都合がよかったため、教育の場や社会の中で訂正することがなかったためと思われます。
5.駆逐説の欠陥を探る
上記のタンポポの個別の性質は、アレロパシー以外は間違ってはいませんでした。一方で、植物や野外の自然を常に見ている人々の間では、セイヨウタンポポ優位の考え方に対する異議が出始めました。端的な事実としては、一つの植木鉢に1本ずつセイヨウタンポポとカントウタンポポを植えておくと、いつまでも両種が生き続けるという反論がありました。風説に用いられたセイヨウタンポポの優位性のうち、アレロパシーは上記のように簡単な事実で否定されました。
では、セイヨウタンポポが持つと考えられた他の性質の優位性はどうでしょうか。1株がつける花(頭状花と呼び、花序、つまり花の集合にあたります)の数、1つの頭状花の中の小さな花(小花と呼び、これが植物学的には花です)の数は一般論としてはセイヨウタンポポの方が大きいのですが、これも年齢や栄養状態で変わります。発芽率も一般論ではセイヨウタンポポの方が高いのですが、光や温度によって状況は色々です。花がよくみられる春に種子を採って播くと、セイヨウタンポポはすぐ一斉に発芽しますが、カントウタンポポはほとんど発芽しません。これだけ見るとセイヨウタンポポの方が圧倒的優位性を持っていると思わされるのですが、カントウタンポポの種子は夏を越して秋になってから一斉発芽します。成長が早い、大きくなるというのも見た目にはそうですが、セイヨウタンポポは順調に育つと発芽の翌春に開花します。一方、カントウタンポポは秋に発芽して、育ちの良いものは翌春に開花します。カントウタンポポの方が半年も早く生殖に入ることができることになります。
種子重はセイヨウタンポポの方が軽いので、より小さな冠毛でも遠くへ飛ぶ可能性があります。カントウタンポポの種子が重いのは、半年の間種子のままで発芽適期を待っている間の栄養を備えているとも考えられます。さらに、カントウタンポポの種子が飛びにくいのは、仲間の近くに種子を落として受精を可能にするには好都合です。セイヨウタンポポのように遠くへ種子を飛ばしたら、花粉を供給してくれる仲間がいないことになります。これらの諸性質はそれぞれのタンポポの生き方の違いであって、もともと比較して優劣をつけるようなものではありません。
6.可哀そうな外来種タンポポ
じつはセイヨウタンポポ優位説には、根本的欠陥がありました。それは種子から芽生えた子どもが1本残らずそのままおとなになるという暗黙の前提がありました。しかもそれに気づいていなかったのです。自然界では、生まれるものは必ず死ぬので、個体が死ぬことを計算に入れていないのは論外でした。当時は、昆虫研究者の間では個体群動態といって1匹1匹の生死を追って、集団の運命を予測する研究が始まっていましたが、植物が死ぬときのことを研究している研究者は病理分野を除いてごく少数で、一般国民には理解がほとんどありませんでした。カントウタンポポの劣性といわれた性質も、種子ができる範囲で綿毛を飛ばし、秋まで待って他の植物が枯れ始めるころに種子から発芽するなど、多くの草本が夏季に旺盛に生育する温暖な日本の田園地帯で個体や集団が生き残るのには適した優れた性質なのです。タンポポは地上に伸びる茎を持ちません(根元から葉が出る)。そのため背が低い植物(背丈は最大で1枚の葉の長さ)ですから、他の植物に覆われると光合成に必要な光を得られず生長できません。
実験的には、東京で春に野外に播いたセイヨウタンポポの種子はすぐ発芽しますが、ほぼ100%秋までに死んでしまいます。梅雨の晴れ間は思いのほか日射が強く地表面が乾燥するし、その後は周りの植物がタンポポの芽生えを覆ってしまいます。それで梅雨明けまでに大半が、秋までにほとんどの個体が死んでしまうのです。これは実験だけでなく、自然に生えたセイヨウタンポポの芽生えでも同じ結果でした。日射が関東ほど強くなく気温が低い北海道や東北地方では、のちに触れる雑種タンポポではない純粋のセイヨウタンポポが見られるのは、発芽後の死亡が本州中部以西より少なくて済むためだろうと推測しています。
7.タンポポ調査の結果から
市民によるタンポポの分布調査によって、どこでもセイヨウタンポポが優勢とは限らないことが明らかになっていきました。大都市の中心地にあっても、例えば大阪城内や皇居周辺には在来種タンポポが見られます。東京都心ではほかに、文京区内の小石川植物園、小石川後楽園、護国寺境内、台東区との境にある旧岩崎邸庭園などで、カントウタンポポが群生し、セイヨウタンポポは少数派でした。また、農地が連続的に残る首都圏の多摩地域では、カントウタンポポが優勢な地区もいくつか見つかりました。大阪では、セイヨウタンポポの優勢な都市の特徴として大規模開発の年代が高度経済成長の時期であることが見えてきました。同様に、東京都と千葉県との県境になっている江戸川の堤防では、整備年代が高度経済成長期を境にして、それ以前の場所にはカントウタンポポが、それ以降の場所ではセイヨウタンポポが優勢であるという高校生による報告もありました。
首都圏ではタンポポ調査にあたって、タンポポの有無にかかわらず調査地点をあらかじめ地図上に一定の距離で決めてそこへ出かけて調べるという方法をとりました。これは、在来種がある場所を見つけたい、調べたいという人間の感情により、在来種生育地が選択的に調査されるのを防ぎ、多人数同時調査の信頼性を増すとともに、出てきた数字を統計的に扱う保障にもなりました。結果として、カントウタンポポは農地や保存されてきた庭園などに集中して出現し、セイヨウタンポポは住宅地や公園、駐車場など都市的土地利用が行われている場所に集中する傾向が明らかとなりました。前者は定期的な土地への人間の作用の下で共存植物が多くみられる環境ですし、後者は人間による一時の大規模な土地かく乱や常に除草などの圧力があって共存植物が少ない環境です。
8.もう一度、タンポポの雑種形成
日本の在来種タンポポと外来種タンポポとの間に雑種ができるということを初めて実験的に証明したのは元新潟大学の森田竜義さんです。彼はヨーロッパのタンポポ研究のメッカであるオランダに留学し、第一人者と言われたステルクさんのもとで実験をしました。ヨーロッパでは、染色体を2セットもったタンポポ(2倍体)と3セット持ったタンポポ(3倍体)は自由に交配して子孫をつくると信じられていました。これらのタンポポは個別の染色体に違いはないので、DNA検査をしても親子関係はわかりません。それで森田さんたちは、ヨーロッパのタンポポとは異なる遺伝情報を持つ日本の2セット染色体を持ったタンポポ(2倍体)を片親にして、染色体の違いからつくる酵素の違いを分析して、親子を決定しました。
そこでは先に触れたように、卵子親は必ず2倍体のタンポポであり。花粉親は3倍体のセイヨウタンポポでも可能という結果が出ました。森田さんはそれまでも、日本で雑種が生まれているということを富士市での調査から気付いていました。のちに同じ分析手法を用いて、愛知教育大学の渡邊幹男さんらが愛知県や大阪府などで雑種タンポポを見つけ、雑種でない純粋のセイヨウタンポポは少ないことをみつけました。さらに、国立農業環境技術研究所(当時の名称)の芝池博幸さんらによって、日本では全国的に雑種タンポポが分布し、純粋のセイヨウタンポポは主として北海道・東北地方で見られ、関東以西では雑種に比べてごく少数派であることがわかってきました。
9.在来種と外来種の識別
それまで、タンポポは頭状花を包む緑の部分(花の集合である花序を包むので、総苞と呼びます。大まかに2層になっていて、内側(上方)のものを内総苞片、外側(下方)のものを外総苞片と呼びます)の形でセイヨウタンポポと在来種タンポポとが区別されてきました。ついでながら、ガクは個別の花を包む皮で、タンポポの場合は綿毛(冠毛)が相当します。外総苞片が下向きに反転するのがセイヨウタンポポ、反転せず上向きになっているのが在来種タンポポとされてきました。元東京学芸大学の小川潔らの研究で、関東地方ではセイヨウタンポポの染色体を3セットと日本のタンポポの染色体を1セット持つ雑種(4倍体雑種)と、それぞれ2セットと1セット持つ雑種(3倍体雑種)があり、4倍体雑種と純粋の3倍体セイヨウタンポポは外総苞片が反転下垂しているものが多く、3倍体雑種はセイヨウタンポポ的なものから、外総苞片が下垂しない在来種的ものまでいろいろな程度の外総苞片の個体が見られること、純粋のセイヨウタンポポ、3倍体雑種、4倍体雑種の中には在来種のように外総苞片が反転下垂しない個体もあることが明らかになり、外総苞片の形で在来種、外来種(あるいは雑種)タンポポの正確な識別はできないことになりました(外総苞片の形の割合の図はこちら)。もっとも、カントウタンポポと、雑種を含むセイヨウタンポポ類は、見た目でも慣れるとある程度識別できます。カントウタンポポが残っている多摩地区では見間違いが少なく、カントウタンポポがほとんど残っていない東京都心部では間違えて雑種をカントウタンポポと判断する率が高く出ました。在来種をみつけたいという人の心情を反映した結果でしょうか。
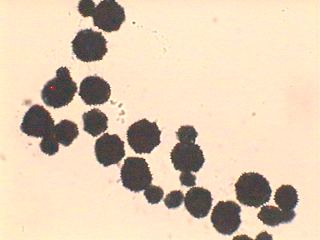
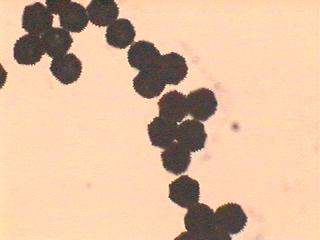
タンポポの在来種、外来種、雑種の識別は、原則としては遺伝子を比較するしかないことになりました。ただ、日本に持ち込まれたセイヨウタンポポが3倍体だったことから、在来種の2倍体種との区別には花粉のサイズが手掛かりになります。本稿のはじめに触れたように、倍数体のタンポポの花粉形成では正常の減数分裂ができないために花粉のサイズがバラバラになります(セイヨウタンポポの花粉の画像はこちら)。一方、2倍体種では正常の減数分裂の結果、花粉のサイズがそろっています(カントウタンポポの花粉の画像はこちら)。それで、花粉サイズがそろっていなければ2倍体ではないと判定できます。100倍程度の携帯顕微鏡で見分けられるので、野外で外総苞片が反転下垂していないタンポポを見つけた場合、2倍体在来種か否かを識別するのに便利です。ただし、日本では在来種の倍数体も広く分布しているので、関東以南九州北部までの平野部でしか使えません。
10.雑種形成の意義
それでは、雑種形成は植物自身にとって、どんな意味があるのでしょうか。セイヨウタンポポは関東以西では純粋の遺伝子構成のままでは残りにくいようです。原産地のヨーロッパでは、例えばロンドンは日本の北海道と同じ緯度にあるように、セイヨウタンポポは比較的冷涼な地域に生えています。加えて放牧もあるため、日本のように夏に草が旺盛に生える環境ではないようです。また、セイヨウタンポポはヨーロッパでは野菜として栽培・食用にされています。
森田さんが日本の2倍体タンポポをヨーロッパに紹介したため、2倍体のセイヨウタンポポ探しがブームとなり、東欧を含めてヨーロッパ各地で2倍体セイヨウタンポポが見つかりました。ただ、ヨーロッパではメジャーなのは3倍体で、2倍体と3倍体のセイヨウタンポポが共存している場所も少なくないとの結果が出ています。
小川らが実験したところ、2倍体のセイヨウタンポポにも、カントウタンポポと同様、自分の花粉では種子ができない自家不和合の性質があります。これは森田さんも独自に確認しています。外国に進出しているのは3倍体セイヨウタンポポだけのようです。セイヨウタンポポと聞くと、アメリカからやって来たと思い込んでいる人がいますが、アメリカ大陸でも山岳地などに在来種タンポポがありますが、都市部では外来種としてセイヨウタンポポが広がっています。たまたま日本には2倍体の在来種タンポポが広範に生育していたので、雑種化が気づかれることになったのでしょう。ただ、日本で見られる雑種タンポポの母親がどの在来種タンポポだったのか、いつどこで雑種が生まれて広がったかはわかっていません。多起源だったと考える方が良いかもしれません。また、標本として残っている頭状花の分析から、1つの遺伝子の差異に注目すると、少なくとも1970年代末には雑種化が起こっていたと推定できるデータもあります。しかし、雑種化を証明するには、複数の遺伝子が同時にチェックされてOKをだす必要があり、古い標本では遺伝子が壊れているため、研究は進んでいません。
ところで、関東以西では純粋の3倍体セイヨウタンポポは少なく、外来種とみられるものの多くが雑種タンポポであることを先に触れました。ここでは、純粋のセイヨウタンポポという植物の見かけの形を捨てて、在来種の頭状花に花粉を送り込み、雑種という形でセイヨウタンポポの遺伝子を温存していると見ることができます。一時期、遺伝子・DNAをテーマにした映画が流行したことがありましたが、セイヨウタンポポではまさに遺伝子こそが主役で、植物という外見は遺伝子を保存する器でしかないというストーリーができます。環境の適しない場所でも、雑種化によってセイヨウタンポポの遺伝子が世界に広がれたのかも知れません。母系のクローン形成も相まって、ホラー映画の主役となれるように思われます。
11.2倍体セイヨウタンポポを日本国内で発見
20年ほど前、南関東地方で緑化資材としてセイヨウタンポポが持ち込まれた例が見つかりました。小川らによって、この集団は3倍体と2倍体のセイヨウタンポポ(画像はこちら)が混合していることが判明しました。この場所の近くには在来2倍体種のタンポポもあり、雑種形成が心配されます。というのは、これまでの雑種は3倍体のセイヨウタンポポを花粉親にして3倍体または4倍体になり、2倍体在来種タンポポ自体の中にセイヨウタンポポの遺伝子は残りません。それで、駆除を徹底させることが理屈の上では可能です。ところが2倍体のセイヨウタンポポは2倍体在来種との間で交配ができ、子孫は2倍体になります。この子孫が在来種と交配を繰り返すと、セイヨウタンポポの遺伝子は薄まっても広く在来種集団の中に行き渡り、外見上は区別がつきにくい可能性が高いので取り除くのが難しくなります。実験的に交配してつくった雑種1代目は外総苞片の一部が下垂反転または内総苞片から剥離するので、なんとか見分けられますが(雑種第1代の画像はこちら)、何代も交配を重ねると見分けができなくなるだろうと推測されます。DNAのチェックなしではお手上げになりそうです。
21世紀のキーワードの一つである生物多様性の保全とは、地域固有の遺伝子構成を守ることが基本です。地域固有の遺伝子構成は、長い時間をかけて環境と生物との相互作用および生物の生き残りや子孫形成の過程での偶然性によって地域ごとに個別に出来上がった歴史的遺産であり、再現できないものです。人間の作用によって、本来出会わないはずの遺伝子を接触させ、自然の作用の結果ではない生き物をつくり、地域固有の遺伝子構成を失わせることは、見かけ上新種形成で種類数が増えたとしても、生物多様性の保全にはならず、むしろその破壊になるという結果をもたらします。だからと言って、人間の意図的・非意図的行為の結果として移動させられた生きものを、外来種ということだけで駆逐・せん滅させるというのも短絡的発想ではないでしょうか。実害が起こらない程度に押さえ込んで、寿命を待つのが対処法でしょう。
セイヨウタンポポは人間の活動をも利用して遺伝子の世界的分布を広げています。また、食材としても栽培されます。2倍体セイヨウタンポポを含む先の南関東の集団は現地ではだんだん衰退していますが、20年たってもまだ健在です。私とタンポポと、どちらが先にあの世に行くか競争ですが、私はこの集団を看取りたいと思っています。
12.もう一度、可哀そうなセイヨウタンポポがひろがったわけ
ところで、セイヨウタンポポと日本の在来タンポポとでは、生き方の違いがあることは紹介しました。個々の性質の大小では、優劣はつけられないことにも触れました。それでもセイヨウタンポポがあちらでもこちらでも見られますし、在来種タンポポが見られる場所は限定されています。これはいったいどういうことなのでしょうか?
カントウタンポポとセイヨウタンポポを実験的に共存させ、除草をせずに人が時々歩くという、昔ながらの田園地域の環境に似た状態をつくって、双方の開花した個体数を調べたことがあります。1年たった春、開花個体数はそれぞれ14と28(比率は1対2)で、 セイヨウタンポポが優位にあるように見えました。ところがスタートから7年たった時点で再調査したところ、それぞれ505と58(約9対1)でした。圧倒的にカントウタンポポが優勢になっていたのです。カントウタンポポが集中して見られる農地や庭園ではこのようなことが起こっていると考えられます。一方、人間の圧力がいつも強く働いている都市的環境では、周りに背が高い草がないので、早く到達できるセイヨウタンポポが生き残れたのでしょう。簡単に言うと、人間の干渉の程度によって、どちらの種類が生き残りやすいかが決まることになります。それも、100とか1000くらいの植物個体数を用いる実験では全滅するので、野外での万あるいは億という単位で生産される種子数の規模でやっと例外的に生き残る個体があってセイヨウタンポポは維持されていると推測されます。多産多死というセイヨウタンポポの性質は、在来種との競争においてではなく、適さない環境の中で生き残る手立てとして再評価できます。新天地である日本の暖温帯域では、過酷な環境下で生きざるを得なかったセイヨウタンポポは、とても可哀そうな存在でした。農業国から工業国への転換、第3次産業のための大都市開発が、セイヨウタンポポに生き残る環境を与えました。そして雑種化という、遺伝子を在来種の子孫のなかに残すことによって日本の暖温帯域に生き残るという戦略をとったという点では、セイヨウタンポポはとてもしたたかな存在と言えるでしょう。
13.東京都心の在来種再点検
東京都心の在来種タンポポであるカントウタンポポについて、ここ50年ほどの変遷を台東区・文京区の地域で振り返ってみましょう。1970年代、両区が接する今の通称「谷根千」地区で見ると、その生育地点は、谷中霊園、寛永寺の徳川将軍墓地の石垣、東京国立博物館庭園、両大師境内、上野駅と鶯谷駅の間の国鉄(今のJR)の土手、国立科学博物館と国立西洋美術館の間(今の西洋博物館新館)の旧凌雲院墓地、不忍池の岸辺、無縁坂、旧岩崎邸庭園、東京大学構内の懐徳館庭園、赤門、農学部と点々と続き、少し飛んで護国寺境内、小石川植物園や小石川後楽園、新江戸川公園(今の細川庭園)がありました。いわば歴史的環境伝いにほぼ連続し、それ以前に広く連続分布していたことをうかがわせるものでした。
カントウタンポポは現在、旧岩崎邸庭園では中央の広場に多数の個体が群生していますが(画像はこちら)、ここが東京都の管理になって庭園として公開されるまで、広場はやや湿っぽく、スゲ類が多く、カントウタンポポは現在のようには多くありませんでした。公開後、多くの入園者がここを歩いたりダンスパーティーを催したので、乾燥化が進んだようですし、背が高い草は生育できなくなりました。ここに周りからカントウタンポポが侵入して大群落になったのでしょう。たぶん、2倍体タンポポの個体ごとの遺伝子に多様性があったので、群落の回復が可能だったと思われます。一方、カントウタンポポは路傍では生育余地がなくなり、墓地、土手、寺院などでは開発のために、不忍池では一時的イベント場設置により次々と失われました。それでもこの地域はまだ、大都市の中心地でありながらカントウタンポポが残っています。寺院や庭園という歴史的文化遺産が、在来種タンポポという自然遺産を残してきたとも言えます。
長い話をお読みいただき、ありがとうございました。内容に関するご質問や、これからタンポポ研究をしてみようという方は著者小川潔までご連絡ください。。
|
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}